根際細菌菌株可提高辣、及花生種子發(fā)芽率,促進幼苗生長
3結果與分析
3.1 3株茶樹根際細菌浸種對辣椒種子萌發(fā)及幼苗生長的影響
利用從茶樹根際分離、篩選到的3株細菌菌株GD 3、KKS-6-N 1及P 5對辣椒進行浸種試驗,萌發(fā)后于盆栽條件下繼續(xù)生長30 d后測定幼苗的相應生長指標。以3株細菌菌液進行了2 h的浸種處理后,辣椒種子的萌發(fā)率均在90%左右,較對照顯著提高了近20%,但不同菌株處理之間并無顯著性差異(p<0.05)。30 d的盆栽實驗表明,與對照相比,GD 3、KKS-6-N 1、P 5浸種處理的辣椒幼苗鮮重分別增加54.55%、63.64%和45.46%,株高分別增長85.14%、75.23%和84.69%,根長分別增長了69.51%、13.17%和129.76%。顯然,3株茶樹根際細菌的浸種處理使辣椒種子的發(fā)芽率明顯提高,且對辣椒的株高和鮮重具有顯著促生作用;其中,P 5浸種處理的植株根長明顯高于對照及其余2個菌株的處理。
3.2 3株茶樹根際細菌浸種對花生種子萌發(fā)的影響
利用3株茶樹根際細菌菌株對花生進行相同的浸種處理,3 d后統(tǒng)計種子的萌發(fā)率(圖1),結果發(fā)現(xiàn),菌劑浸種處理組的發(fā)芽率均在90%以上,較對照種子的發(fā)芽率提高了20%以上,3株細菌菌株對種子的萌發(fā)具有明顯的促進效果,但不同菌株處理之間對種子萌發(fā)率并無顯著差異(圖1)。
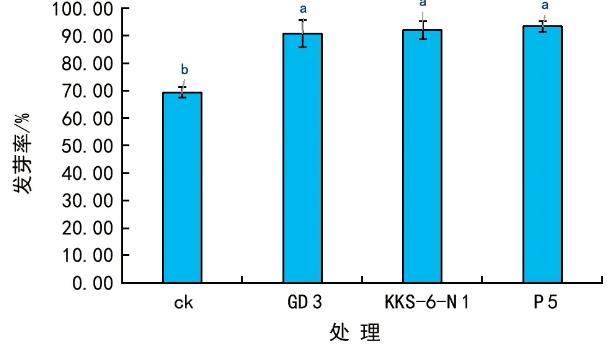
圖1 3株茶樹根際細菌對花生種子萌發(fā)的影響
3.3 3株茶樹根際細菌灌根處理對花生幼苗生長的影響
為了進一步探討根際細菌對花生幼苗生長的影響作用及內(nèi)在機制,分別利用3株菌液進行灌根處理,培育30 d后測定花生的生長性狀(表2),發(fā)現(xiàn)3株菌株對花生幼苗生長的影響呈現(xiàn)一定差異。與對照相比,GD 3、KKS-6-N 1、P 5灌根處理的花生幼苗鮮重分別增長4.63%、26.83%和35.12%,株高分別增加了5.99%、16.22%和32.73%,根重分別增長21.62%、29.73%和35.14%。生理指標的測定結果顯示,GD 3和KKS-6-N 1接種組的葉綠素含量略低于對照,而P 5接種組的葉綠素含量較對照提高了13.47%。結果(圖2)表明,P 5菌株的灌根處理對花生幼苗的促生效果顯著,其次為KKS-6-N 1。
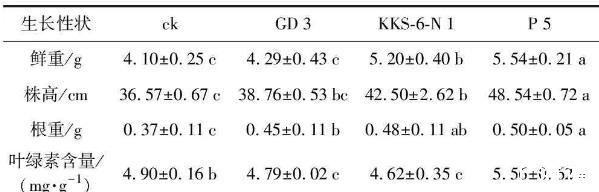
表2 3株茶樹根際細菌對花生幼苗生長的影響
3.4 P 5菌株灌根處理對花生植株營養(yǎng)元素的影響
由于P 5菌株可以顯著促進花生幼苗鮮重和株高的增加,因而經(jīng)P 5灌根處理的花生植株進行了營養(yǎng)指標測定(表3),發(fā)現(xiàn)接種組植株的全氮增高了13.56%,與對照呈現(xiàn)顯著差異;全鉀含量略有提升,但差異不明顯,而全磷含量基本無改變(p<0.05)。

注:圖左3株花生為P 5菌株處理,圖右3株為ck。圖2根際促生菌菌株P 5對花生幼苗生長的影響
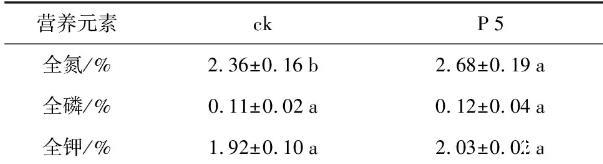
表3根際促生菌菌株P 5對花生植株營養(yǎng)元素的影響
3.5 P 5菌株灌根處理對花生根際土壤三大菌群和功能菌群數(shù)量的影響
由于土壤微生物在土壤營養(yǎng)物質(zhì)循環(huán)及維持土壤肥力方面起著重要作用,實驗擬進一步從土壤菌群數(shù)量和土壤酶活入手,解析該菌株對花生的促生機制。對P 5菌株灌根處理的花生根際土壤及對照土壤進行三大菌群總數(shù)的測定(表4),結果發(fā)現(xiàn),處理組根際土壤中的細菌總數(shù)較對照增長了2.63倍,真菌總數(shù)增長了2倍,細菌數(shù)/真菌數(shù)之比增加了1.32倍,菌劑處理使得細菌和真菌數(shù)量較對照呈現(xiàn)顯著差異,而放線菌總數(shù)并無明顯差異(p<0.05)。由于P 5處理后的花生根際土壤中細菌總數(shù)顯著提高,因而對根際土壤中與促生相關的功能菌群數(shù)量進行了測定(表4),發(fā)現(xiàn)氨化細菌及固氮菌數(shù)分別較對照增高了2倍和2.44倍,溶磷菌數(shù)增高了4.07倍,與土壤氮磷含量直接相關的氨化細菌、固氮菌及溶磷菌數(shù)均明顯高于對照,而解鉀菌數(shù)略有下降,但差異不顯著(p<0.05)。
3.6 P 5菌株灌根處理對花生根際土壤酶活性的影響
酶活性的測定結果(表5)顯示,與對照相比,處理組根際土壤的蔗糖酶活性增加了1.41倍,過氧化氫酶活性增長了2.65倍,脲酶增長了2.33倍,中性磷酸酶活性基本無改變,表明P 5浸種處理的花生根際土壤中,蔗糖酶、過氧化氫酶及脲酶酶活性均較對照有顯著提高(p<0.05)。
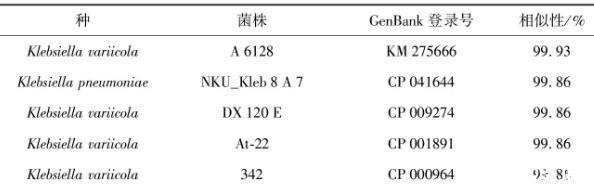
表6 P 5菌株的16 S rRNA基因相似性比對
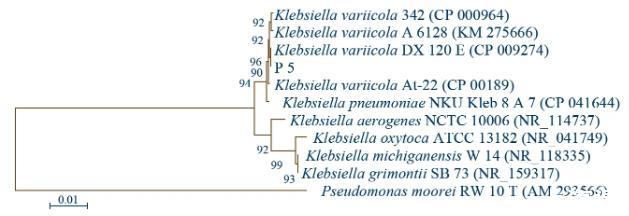
圖3 P 5菌株的16 S rRNA基因系統(tǒng)發(fā)育樹
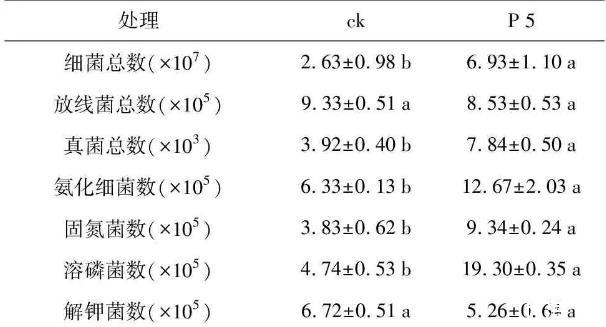
表4根際促生菌菌株P 5對花生根際土壤三大菌群數(shù)量(cfu·g-1)的影響
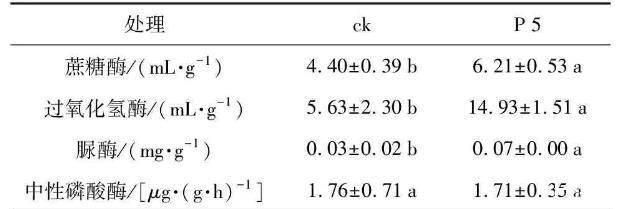
表5根際促生菌菌株P 5對花生根際土壤酶活性的影響
3.7 P 5菌株的分子鑒定及系統(tǒng)發(fā)育樹的構建
利用細菌16 S rRNA基因的通用引物對27 F/1492 R對P 5細菌的總DNA進行PCR擴增,測序結果獲得1 405 bp的核苷酸序列,經(jīng)NCBI網(wǎng)站上進行BLAST比對,發(fā)現(xiàn)與其一致性最高的菌株為克雷伯氏菌屬、變棲克雷伯氏菌(Klebsiellavariicola),與該種A 6128菌株的16 S rRNA基因相似性為99.93%,此外與多株變棲克雷伯氏菌菌株及肺炎克雷伯氏菌NKU_Kleb 8 A 7菌株的相似性均為99.86%(表6)。與同屬內(nèi)的不同種構建系統(tǒng)發(fā)育樹(圖3),可見P 5菌株與Klebsiellavariicola的不同菌株聚于一大枝,且與該種DX 120 E菌株(CP 009274)的16 S rRNA基因在系統(tǒng)樹上相鄰,因而結合分子鑒定及系統(tǒng)發(fā)育樹,可將P 5菌株鑒定為Klebsiellasp.。
4結論與討論
本研究中使用的3株細菌菌株GD 3、KKS-6-N 1和P 5均是從茶樹根際分離篩選獲得的,浸種試驗表明,3株細菌均可使辣椒及花生種子的發(fā)芽率達到90%以上,較對照相比,發(fā)芽率可提高20%左右,對辣椒及花生種子的萌發(fā)均具有顯著的促進作用。在農(nóng)業(yè)生產(chǎn)上,目前的花生栽培通常是種子經(jīng)拌種或包衣后進行播種,利用菌劑進行種子包衣處理不失為一個提高種子萌發(fā)的有效方式。而辣椒種子常因成熟度不夠、采種技術不當或貯藏方法等原因造成發(fā)芽率降低[22],研究結果為利用3株細菌菌劑進行拌種或包衣處理奠定了基礎。
但是,促進種子萌發(fā)的菌株不一定會對植株的生長有顯著促進作用。張振建等報道,烏拉爾桿菌PN 133和土壤根瘤桿菌Y 42的浸種均可使莧菜和油麥菜的發(fā)芽率提高,而灌根處理則促生效果并不明顯[23]。利用2株溶磷菌進行青稞種子的萌發(fā)試驗中,菌株10 BN-11對增加青稞發(fā)芽率及促進生長效果明顯,菌株12-BN-6對株高、根長和鮮重具有明顯的促進作用,但可抑制種子的萌發(fā)[24]。王艷燕等的研究發(fā)現(xiàn),在9種可促進辣椒發(fā)芽率提高的菌株中,后續(xù)的穴盤育苗有促生效果的僅有4株菌,其中對辣椒下胚軸和主根長度影響顯著的GH 6-1和GH 5-3并未在苗期表現(xiàn)出顯著的促生作用,說明有些菌雖能促進種子萌發(fā),但未必能在后期幼苗生長過程中促進其對養(yǎng)分的吸收[25]。本研究的浸種實驗雖顯示3株細菌均可明顯提高辣椒和花生種子的發(fā)芽率,且不同菌株的處理并無顯著差異;但辣椒種子萌發(fā)后的生長過程中,3株細菌對其后續(xù)的影響略有不一致;而且,在利用菌液的灌根處理中,發(fā)現(xiàn)P 5菌株對花生幼苗的促生效果顯著高于KKS-6-N 1,GD 3對幼苗株高及鮮重的影響則并不明顯,這也與報道的相一致。因而對于根際細菌促生作用的研究中,有必要分別對種子萌發(fā)和幼苗生長進行研究,綜合評判根際細菌菌株的促生作用。
研究顯示,根際促生菌可提高種子的發(fā)芽率和芽長,進而提高植株的生物量,但植物對所分離的功能菌株是否具有根際效應,是菌株發(fā)揮功能的關鍵[26]。康貽軍等在評價PseudomonaschlororaphisRA 6和BacilluspumilusWP 8浸種和拌土對豇豆生長的作用時,發(fā)現(xiàn)菌液的浸種處理及108cfu·g-1劑量的菌株拌土處理均可顯著提高豇豆的株高,認為浸種處理從原理上講,應該不會對非根際土壤產(chǎn)生較大影響[27]。Probanza等研究表明,PGPR對植物土壤中土著微生物數(shù)量及群落結構的影響可能是其促生的另一個機制[28]。研究表明,土壤酶活性與土壤速效養(yǎng)分密切相關,而土壤微生物數(shù)量大、繁殖快、活性強,直接影響到植物根際有機質(zhì)的分解和養(yǎng)分的轉(zhuǎn)化[29]。因而,為了進一步解析和探究P 5菌株對花生幼苗生長的促進機制,本研究對接種P 5菌株的花生根際土壤進行了土壤微生物及酶活的測定。由于細菌、放線菌和真菌是土壤微生物的三大類群,構成了土壤微生物的主要生物量,它們的區(qū)系組成和數(shù)量變化能反映土壤生物活性水平[30]。
研究結果顯示,P 5處理組根際土壤中的細菌總數(shù)較對照增長了2.63倍,真菌總數(shù)增長了2倍,細菌總數(shù)/真菌總數(shù)之比增加了1.32倍,而放線菌總數(shù)并無明顯差異(p<0.05)。多數(shù)報道表明,接種促生菌均可不同程度地提高土壤細菌總數(shù)和放線菌數(shù),降低真菌數(shù)量[31-33]。王興祥等研究認為,細菌型土壤是土壤肥力提高的一個生物指標,真菌型土壤是地力衰竭的標志[34]。但在巨尾桉接種固氮菌或解鉀菌的土壤中,發(fā)現(xiàn)細菌及放線菌數(shù)量增多,而真菌數(shù)量均不同程度的增高[35]。在研究中,真菌數(shù)量雖有明顯提高,但細菌數(shù)/真菌數(shù)之比仍高于對照,接種組土壤仍表現(xiàn)為細菌型土壤。土壤酶活性在一定程度上可以反映土壤肥力狀況。土壤蔗糖酶活性強弱能反映土壤的熟化程度和肥力水平[36],過氧化氫酶活性與土壤呼吸作用及土壤微生物活動密切相關[37],脲酶活性可在一定水平上反映土壤的供氮水平與能力[38],磷酸酶在土壤有機磷向無機磷的分解轉(zhuǎn)化方面起著重要作用[39]。
研究顯示,除中性磷酸酶外,P 5接種組的根際土壤蔗糖酶、脲酶及過氧化氫酶均顯著高于對照,表明根際細菌P 5的處理活躍了土壤微生物;對功能菌群數(shù)量的進一步測定結果表明,氨化細菌數(shù)、固氮菌數(shù)及溶磷菌數(shù)顯著增高(p<0.05)。氨化細菌及固氮菌數(shù)量的顯著提高可以提供植物更多所需的氨氮,接種組植株全氮含量的明顯提高與土壤脲酶等酶活性的增高、氨化細菌及固氮菌數(shù)的顯著提升相一致,這顯然是促進花生生長的原因之一。
多項研究也已證實土壤酶活性與大豆、小麥及玉米產(chǎn)量呈顯著正相關關系[40-41]。此外,P 5菌株經(jīng)分子鑒定為克雷伯氏菌屬,與KlebsiellavariicolaDX 120 E的同源性最高;而Klebsiellavariicola是2004年才發(fā)現(xiàn)的新種,作為內(nèi)生菌,并由于其優(yōu)良的固氮性能被報道[42-43];本研究中的P 5菌株是具有一定固氮能力的茶樹根際細菌,因而接種了克雷伯氏菌屬的P 5菌株后,顯著提升了花生的全氮含量。值得注意的是,P 5菌株處理的花生全磷含量與對照基本無差異,這與根際土壤中性磷酸酶活無顯著變化相一致,而溶磷菌數(shù)明顯增高的原因還有待進一步研究。
綜上所述,Klebsiellasp.P 5菌株的浸種處理可以顯著促進辣椒和花生種子的萌發(fā),灌根處理可以顯著促進花生幼苗的生長,植株的全氮含量明顯提高,這與根際土壤氨化細菌及固氮菌數(shù)量的顯著增多,土壤蔗糖酶、脲酶及過氧化氫酶活性的顯著增高直接相關,研究結果為下一步利用P 5菌株進行拌種或田間施用奠定了理論基礎。
相關新聞推薦
1、呼寧散對雞肺源大腸桿菌生長曲線、細胞壁的影響及抑制效果——摘要