微生物合成食品功能因子方法、研究策略和進(jìn)展(二)
2重編程細(xì)胞能量代謝
能量代謝在細(xì)胞生長和中心代謝途徑中發(fā)揮重要作用,操縱能量代謝是優(yōu)化微生物生長適應(yīng)性和實(shí)現(xiàn)產(chǎn)物高產(chǎn)量、高產(chǎn)率和高生產(chǎn)強(qiáng)度相統(tǒng)一的有效策略[65,66].ATP作為最重要的能量來源,調(diào)控ATP供應(yīng)是調(diào)節(jié)細(xì)胞工廠代謝流分布、生物量積累和產(chǎn)物合成的重要手段[67,68].有氧條件下細(xì)胞主要通過氧化磷酸化途徑合成ATP,因此操縱該途徑可以有效調(diào)節(jié)細(xì)胞內(nèi)ATP的濃度,其中主要通過調(diào)控NADH水平、ATP合酶活性和電子傳遞鏈來調(diào)節(jié)ATP代謝.
2.1調(diào)控NADH供給
細(xì)胞內(nèi)NADH主要來源于糖酵解、脂肪酸氧化和三羧酸循環(huán),所以可以根據(jù)NADH代謝來調(diào)控NADH相關(guān)酶的活性進(jìn)而調(diào)節(jié)胞內(nèi)ATP水平.Zhu和Shimi zu[69]敲除E.coli中編碼丙酮酸甲酸裂解酶的pfl基因,從而導(dǎo)致胞內(nèi)NADH/NAD+比率顯著升高,ATP/AMP水平降低,同時(shí)在以葡萄糖為碳源時(shí)菌株的糖酵解通量顯著增加.研究人員通過調(diào)控E.coli TCA循環(huán)中相關(guān)基因的表達(dá),增強(qiáng)了TCA循環(huán)的碳通量和胞內(nèi)ATP水平,導(dǎo)致β-胡蘿卜素[70]和番茄紅素[71]產(chǎn)量顯著增加.為實(shí)現(xiàn)異丁醇的快速高效生產(chǎn),研究人員對合成異丁醇的B.subtilis進(jìn)行代謝網(wǎng)絡(luò)分析,揭示了NADPH對異丁醇生物合成的重要性[72].因此在氧化還原失衡且NADPH過量的情況下,強(qiáng)化表達(dá)來源于E.coli的udhA基因提高胞內(nèi)NADH和ATP濃度,最終細(xì)胞生長性能得到優(yōu)化,異丁醇產(chǎn)量提高.敲除S.cerevisiae中消耗NADH的乙醇脫氫酶的同時(shí)過表達(dá)生成NADH的乙醛脫氫酶,可提高胞內(nèi)NADH的水平和甘油的產(chǎn)量[73].此外,在培養(yǎng)基中補(bǔ)充依賴NAD+脫氫酶的相關(guān)底物[74],可以有效調(diào)控胞內(nèi)ATP濃度,從而提高產(chǎn)物合成效率.通過調(diào)控NADH供給實(shí)現(xiàn)上調(diào)或下調(diào)胞內(nèi)ATP水平的策略較為清晰簡便,且調(diào)控效果顯著,但NADH濃度的變化可能會破壞胞內(nèi)氧化還原的平衡,因此需要考慮細(xì)胞的氧化還原狀態(tài)是否利于產(chǎn)物合成.
2.2調(diào)控F0F 1-ATP合酶表達(dá)
在氧化磷酸化過程中,電子傳遞鏈產(chǎn)生的質(zhì)子動力通過ATP合酶驅(qū)動ATP生成,因此操縱F0F 1-ATP合酶表達(dá)是調(diào)控ATP水平最直接有效的方法.在E.coli中強(qiáng)化ATP合酶基因操縱子的表達(dá)提升胞內(nèi)ATP水平,可有效提高β-胡蘿卜素的產(chǎn)量[70].Zhang等人[75]在S.cer evisiae中表達(dá)來源于擬南芥(Arabidopsis thaliana)的ATP6基因,提高了ATP水平;后將該基因在產(chǎn)朊假絲酵母(Candida utilis)中進(jìn)行表達(dá),提高了胞內(nèi)F0F 1-ATP合酶的活性和胞內(nèi)ATP水平,同時(shí)菌株的生物量提高且S-腺苷甲硫氨酸和谷胱甘肽的合成增強(qiáng)[76].
由于ATP合酶的F1亞基具有水解ATP的功能,因此表達(dá)該亞基會促進(jìn)ATP水解從而降低ATP的濃度.在E.coli中表達(dá)ATP合酶的F1亞基降低了胞內(nèi)ATP水平,并顯著提高菌株對葡萄糖的攝取速率和乳酸合成能力[77].利用類似的策略,在糖多孢紅霉菌(Saccharopo lyspora erythraea)表達(dá)F1亞基導(dǎo)致較低的[ATP]/[ADP]比,維持了細(xì)胞的氧化還原平衡狀態(tài),紅霉素合成能力提高28%[78],同樣該策略提高了乳酸乳球菌(Lactococ cus lactis)的乙偶姻合成能力[79].因此對于可產(chǎn)生質(zhì)子動力的菌株,操縱ATP合酶活性可有效調(diào)控胞內(nèi)ATP的供應(yīng),并改善生產(chǎn)菌株生物量的累積和與ATP合成相關(guān)的目標(biāo)代謝物的生產(chǎn).
2.3調(diào)控電子傳遞鏈
在氧化還原反應(yīng)中,電子傳遞鏈上電子轉(zhuǎn)移過程中產(chǎn)生的質(zhì)子梯度可用于胞內(nèi)ATP合成.在有氧發(fā)酵中,氧氣是電子傳遞鏈的最終電子受體,因此氧氣濃度是影響胞內(nèi)ATP含量的關(guān)鍵因素[80].尤其是某些高密度、高能量的發(fā)酵體系中氧氣的豐度是ATP合成的重要條件[81].調(diào)控發(fā)酵罐的攪拌速度或在發(fā)酵過程通入氧氣是較為簡單的氧氣供給策略[82].另一種策略是添加一定濃度的正己烷、正庚烷、血紅蛋白和正十六烷等攜氧載體,該策略也可提高氧濃度[83~86].E.coli發(fā)酵過程中血紅蛋白的添加使細(xì)胞生物量提高約11%[85].Li等人[86]在培養(yǎng)基中加入正十六烷,促進(jìn)S.cerevisiae胞內(nèi)ATP積累,S-腺苷甲硫氨酸的產(chǎn)量達(dá)到2.27 g/L.此外,調(diào)控pH維持在酸性條件下可以提高胞內(nèi)ATP水平.因?yàn)檩^低pH產(chǎn)生的質(zhì)子梯度可驅(qū)動呼吸鏈中的F0F 1-ATP合酶活性[87,88].雖然通過調(diào)控電子傳遞鏈可以調(diào)節(jié)ATP代謝,但由于其涉及多個(gè)基因的調(diào)控,難以通過簡單的單基因或幾個(gè)相關(guān)基因的調(diào)控來調(diào)節(jié)胞內(nèi)ATP水平.
ATP作為一類重要的輔因子,對代謝反應(yīng)的速率、細(xì)胞的生長和產(chǎn)物合成具有重要的調(diào)控意義.胞內(nèi)ATP水平對不同產(chǎn)物的影響不同,胞內(nèi)高濃度的ATP可以促進(jìn)多糖等食品功能因子的合成[89],而谷氨酸等代謝物的合成則需要維持低濃度ATP,因此需要平衡宿主胞內(nèi)ATP水平和細(xì)胞生長之間的關(guān)系,從而實(shí)現(xiàn)產(chǎn)品的高效合成.
3細(xì)胞生長與產(chǎn)物合成偶聯(lián)
采用傳統(tǒng)微生物代謝工程策略對細(xì)胞工廠進(jìn)行改造可能會導(dǎo)致代謝通量失衡或代謝負(fù)擔(dān),從而使得細(xì)胞生長性能受損和生產(chǎn)性能減弱,特別是具有高度工程化或遺傳回路的微生物往往會在長期發(fā)酵中失去生產(chǎn)表型[90,91].為了解決該問題,一種策略是將細(xì)胞生長與產(chǎn)物合成相偶聯(lián),使生產(chǎn)適應(yīng)細(xì)胞的生長,實(shí)現(xiàn)生長和產(chǎn)物合成之間的平衡[92,93].
3.1營養(yǎng)缺陷系統(tǒng)與產(chǎn)物合成偶聯(lián)
營養(yǎng)缺陷與產(chǎn)物合成進(jìn)行偶聯(lián)是一種生長與生產(chǎn)偶聯(lián)的方法,其中產(chǎn)物合成對于細(xì)胞生長是必需的.Atsumi和Liao[94]在異亮氨酸營養(yǎng)缺陷型E.coli中引入檸蘋酸途徑合成α-酮戊二酸,從而為1-丙醇和1-丁醇的合成提供前體,且α-酮戊二酸可進(jìn)一步轉(zhuǎn)化為異亮氨酸,使得該菌株能夠恢復(fù)生長能力,但生長速率仍低于直接添加異亮氨酸的情況.后對該途徑的關(guān)鍵酶CimA進(jìn)行定向進(jìn)化,篩選得到生長速率高于野生型的突變株,并顯著提高1-丙醇和1-丁醇產(chǎn)量.Ye等人[95]發(fā)現(xiàn),合成番茄紅素的S.cerevisiae在繼代培養(yǎng)時(shí)出現(xiàn)白色菌落,猜測可能是由于GAL調(diào)控系統(tǒng)的破壞導(dǎo)致途徑基因轉(zhuǎn)錄被抑制從而不能合成番茄紅素,所以出現(xiàn)菌株表型退化的現(xiàn)象.菌株是需要表達(dá)URA3才能正常生長的尿嘧啶缺陷型菌株,編碼番茄紅素合成的基因由GAL啟動子驅(qū)動表達(dá).為了實(shí)現(xiàn)生長與合成偶聯(lián),研究人員利用GAL啟動子同時(shí)控制URA3和番茄紅素途徑基因的表達(dá),從而導(dǎo)致在長期培養(yǎng)中GAL系統(tǒng)破壞的菌株無法正常生長.當(dāng)葡萄糖存在時(shí),葡萄糖誘導(dǎo)的啟動子PH XT1開啟URA3的表達(dá),細(xì)胞正常生長.當(dāng)葡萄糖耗盡時(shí),PG AL啟動子激活產(chǎn)物合成相關(guān)基因和偶聯(lián)的URA3基因表達(dá),從而賦予生產(chǎn)細(xì)胞生長優(yōu)勢.將該策略用于S.cerevisiae高效合成(+)-瓦倫亞烯,產(chǎn)量達(dá)到16.6 g/L.
此外,研究人員拓展了模塊化細(xì)胞(modular cell,MODCELL)[96]的設(shè)計(jì)原理以優(yōu)化生長偶聯(lián)效果.模塊化細(xì)胞是一種營養(yǎng)缺陷型細(xì)胞,包含必要但不足以支持細(xì)胞生長和維持代謝的中心代謝途徑,因此必須與可變的產(chǎn)物合成模塊偶聯(lián).該模塊可以輔助細(xì)胞生長,維持細(xì)胞代謝并增強(qiáng)產(chǎn)物的合成,通過優(yōu)化兩者之間的偶聯(lián)強(qiáng)度可以快速篩選最優(yōu)生產(chǎn)宿主,并通過乙醇的生物合成進(jìn)行了驗(yàn)證[97].
3.2基于群體質(zhì)量控制和代謝產(chǎn)物成癮系統(tǒng)的生長與產(chǎn)物合成偶聯(lián)
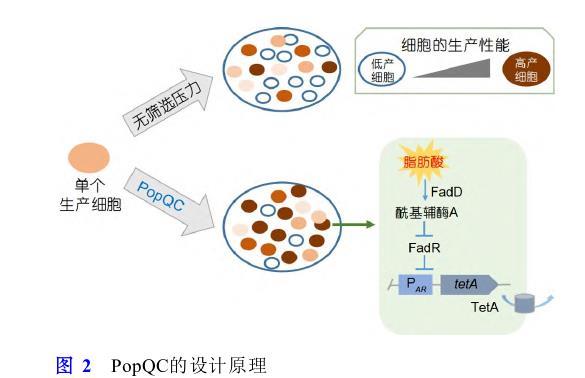
代謝負(fù)擔(dān)常常導(dǎo)致生產(chǎn)細(xì)胞表現(xiàn)出降低的生長適應(yīng)性,由于遺傳和不可遺傳變異導(dǎo)致的非生產(chǎn)細(xì)胞亞群由于缺乏代謝負(fù)擔(dān)而恢復(fù)生長適應(yīng)性,并可以快速占據(jù)細(xì)胞群體,從而降低生產(chǎn)能力[98,99].對抗代謝異質(zhì)性的一個(gè)策略是將代謝產(chǎn)物的合成與細(xì)胞生長進(jìn)行偶聯(lián),從而解決非生產(chǎn)亞群導(dǎo)致的細(xì)胞群體生產(chǎn)能力流失的問題.對此,Xiao等人[100]提出了群體質(zhì)量控制系統(tǒng)(population quality control,PopQC)的概念.首先在E.coli中開發(fā)一種基于調(diào)控因子FadR且響應(yīng)目標(biāo)產(chǎn)物的生物傳感器,該傳感器響應(yīng)游離脂肪酸的濃度,在低產(chǎn)細(xì)胞或者非生產(chǎn)細(xì)胞中轉(zhuǎn)錄調(diào)控因子FadR與PA R啟動子結(jié)合,抑制編碼四環(huán)素外排蛋白基因tetA的表達(dá);而在高產(chǎn)細(xì)胞內(nèi),隨著游離脂肪酸濃度的增加,其會解除FadR對相應(yīng)啟動子的阻遏作用,tetA基因正常表達(dá).因此,在四環(huán)素存在的情況下,PopQC富集高產(chǎn)細(xì)胞并使之占據(jù)主導(dǎo)地位,最終游離脂肪酸的產(chǎn)量提高3倍(圖2),相同的策略使得酪氨酸產(chǎn)量提高2.6倍.同樣,Rugbjerg等人[101]利用胞內(nèi)必需過程,
E.coli中設(shè)計(jì)了正向響應(yīng)產(chǎn)物MVA的遺傳回路以調(diào)控必需基因的表達(dá).當(dāng)MVA產(chǎn)量較高時(shí),生物傳感器與MVA結(jié)合從而激活必需基因的轉(zhuǎn)錄表達(dá),導(dǎo)致菌株具有較高的生長速率;而當(dāng)不存在MVA時(shí),必需基因的表達(dá)也會被限制,從而導(dǎo)致較低的生長速率.因此在整個(gè)細(xì)胞群體中,生長緩慢的非生產(chǎn)細(xì)胞比例逐漸減少,具有高生長速率生產(chǎn)細(xì)胞逐漸占主導(dǎo).最終,未改造的菌株在經(jīng)過50次傳代后MVA合成能力逐漸喪失,而引入產(chǎn)物成癮系統(tǒng)的菌株在傳代95次后仍然擁有高產(chǎn)能力,并且該系統(tǒng)無需使用抗生素并且不存在宿主受限的問題.研究人員還將產(chǎn)物柚皮素成癮與負(fù)向自調(diào)控系統(tǒng)相結(jié)合,使得成癮酵母細(xì)胞培養(yǎng)324代后仍擁有90.9%的柚皮素生產(chǎn)能力,而正常細(xì)胞在傳代300代后丟失94.5%的生產(chǎn)能力,且恢復(fù)了正常生長[102].該策略同樣適用于以E.coli為底盤細(xì)胞合成柚皮素的研究[103].
代謝異質(zhì)性已成為影響細(xì)胞生產(chǎn)性能的主要問題,通過激勵(lì)生產(chǎn)細(xì)胞和懲罰非生產(chǎn)細(xì)胞的方式可以用于對抗這種限制,從而通過富集生產(chǎn)細(xì)胞亞群穩(wěn)定維持生產(chǎn)表型。
食品功能因子的可持續(xù)生物合成面臨的一個(gè)主要挑戰(zhàn)是擴(kuò)大生產(chǎn)規(guī)模并維持高產(chǎn)性能.但是,異源基因表達(dá)引起的代謝負(fù)擔(dān)會產(chǎn)生選擇性壓力,從而導(dǎo)致培養(yǎng)體系中非生產(chǎn)種群的自發(fā)形成,使得高生產(chǎn)能力但適應(yīng)性較低的細(xì)胞喪失優(yōu)勢,最終導(dǎo)致整體生產(chǎn)性能下降.因此,將細(xì)胞生長與產(chǎn)物合成偶聯(lián)可以很好地解決這類矛盾,從而提升細(xì)胞群體的生產(chǎn)性能.
相關(guān)新聞推薦
1、反芻動物瘤胃菌群組成和數(shù)量、影響因素及與代謝性疾病關(guān)系